Caracol es el nombre común de los moluscos gasterópodos
provistos de una concha espiral. Hay caracoles marinos (a veces denominados
caracolas), dulceacuícolas y terrestres. Son muy apreciados por el hombre,
tanto con fines coleccionistas como gastronómicos.

| Caracol romano, Helix pomatia | |
| Clasificación científica | |
|---|---|
| Reino: | Animalia |
| Filo: | Mollusca |
| Clase: | Gastropoda |
| Subclase: | Orthogastropoda |
| Superorden: | Heterobranchia |
| Orden: | Pulmonata |
CARACTERISTICAS:
Los caracoles se mueven con lentitud alternando
contracciones y elongaciones de su cuerpo. Producen mucus para ayudarse en la
locomoción reduciendo la fricción y permitiéndoles el desplazamiento por zonas
de elevada pendiente debido a la untuosidad del mismo. Esta mucosidad
contribuye a su regulación térmica; también reduce el riesgo del caracol ante
las heridas y las agresiones externas, principalmente bacterianas y fúngicas, y
los ayuda a ahuyentar insectos potencialmente peligrosos como las hormigas. El mucus
sirve además al caracol para desprenderse de ciertas sustancias tóxicas como
los metales pesados.
Cuando se retraen en su concha, segregan un tipo especial
de mucosidad para cubrir la entrada que al solidificarse forma una estructura
llamada opérculo. El opérculo de algunos caracoles tiene un olor agradable
cuando se quema, por eso a veces se usa como un constituyente del incienso.

Anatomía de un caracol de jardín. 1: concha 2: hígado 3:
pulmón 4: ano 5: poro respiratorio 6: ojo 7: tentáculo 8: ganglios cerebrales
9: conducto salival 10: boca11: buche 12: glándula salival 13: poro genital 14:
pene 15: vagina 16: glándula mucosa 17: oviducto 18: saco de dardos 19: pie 20:
estómago 21: riñón 22: manto23: corazón 24: vasos deferentes.
En invierno o en estaciones secas, muchas especies
terrestres o de agua dulce hibernan en su concha sellándose con el opérculo,
que les sirve de protección para la hibernación y que se destruye en la
primavera o cuando el entorno se hace más húmedo. Algunas especies se reúnen e
hibernan en grupos mientras que otras se entierran antes de la hibernación.
El caracol de tierra más grande es el caracol gigante
africano (Achatina fulica) que puede medir hasta 30 cm; Pomacea maculata
(familia Ampullariidae), el caracol de manzana gigante es el caracol más grande
de agua dulce, con un tamaño que alcanza los 15 cm de diámetro y más de 600 g
de peso. El más grande de todos los caracoles es el Syrinx aruanus, una especie
marina que vive en Australia.
La concha del caracol es un ejemplo de la representación
de Phi en la naturaleza. Las formas de las conchas de ciertos caracoles de mar
(Conus, Cymbiola) están constituidas por la versión biológica de un autómata
celular.
Cuando el caracol crece, también lo hace su concha. Un
caracol cerrará una sección de su concha y añadirá una nueva cámara al crecer,
cada cámara será más grande que la anterior por un factor constante. Como
resultado, la concha formará una espiral logarítmica. En algún momento, el
caracol construye un reborde alrededor de la abertura de la concha, deja de
crecer, y comienza a reproducirse.
La concha del caracol y las cubiertas de los huevos están
formadas principalmente por carbonato de calcio como las conchas de otros
moluscos. A causa de esto, requieren una buena cantidad de calcio en su dieta y
ambiente acuoso para producir una concha fuerte. Una carencia de calcio, o una
fluctuación en el nivel de pH en su entorno, probablemente hará que su concha
sea fina, se raje, o tenga agujeros. Por lo general, un caracol puede reparar
su daño en la concha con el tiempo, si sus condiciones de vida mejoran, pero
algún deterioro lo bastante grave podría ser fatal para el caracol. Por eso los
caracoles se desarrollan mejor en las zonas calizas. Donde el carbonato cálcico
escasea, algunas especies faltan y otras, las más adaptables, tragan
piedrecitas que contienen calcio, roen huesos, pintura caliza o plantas ricas
en calcio.
Segmentación:
Los caracoles se caracterizan por presentar una
segmentación holoblástica espiral al igual que la mayoría de moluscos, este
tipo de segmentación puede ser diferenciada fácilmente de la segmentación
radial, debido a que el plano de segmentación no es paralelo o perpendicular al
eje animal-vegetal del cigoto, en cambio el plano es oblicuo, lo que lleva a la
disposición en espiral de las blastómeras durante los primeros estadios del
desarrollo; además este plano de división permite que haya mayor área de
contacto entre las células llevando así a un estado termodinámicamente más
estable. Una de las principales características de los animales que presentan
este tipo de segmentación es el número limitado de divisiones celulares previas
a la gastrulación, esto permite llevar estudios de forma muy específica para
determinar el mapa de destino de cada célula de la blástula. A diferencia de
otros animales, en los caracoles no se da la formación de un blastocele en la
blástula y esta es conocida estereoblástula, Durante las primeras dos
segmentaciones, las cuales se dan casi meridionalmente, se producen cuatro macrómera
de gran tamaño, siendo la macrómera D la más grande (el tamaño entre
blastómeras varía según la especie). Durante las siguientes divisiones, se van
a formar micrómeras a partir del polo animal de cada macrómera. Cada uno de los
cuartetos de micrómeras resultantes puede ubicarse a la derecha o a la
izquierda de su macrómera correspondiente creando así el patrón espiral.
Segmentación espiral del caracol.
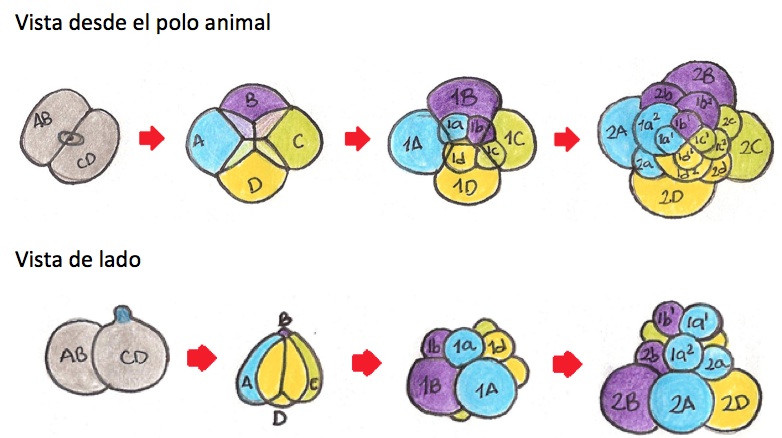
Segmentación espiral del caracol
Plano
de segmentación:
La determinación
del plano de segmentación (derecha o izquierda) se encuentra controlada por
factores citoplasmáticos que se encuentran al interior del ovocito. En los
caracoles, las vueltas en espiral de las conchas pueden ser dextrales
(abriéndose hacia la derecha), o sinistrales (izquierda), esto se encuentra
determinado por un solo par de genes, sin embargo el genotipo del caracol en
desarrollo no determina la orientación de la concha, ésta se encuentra
determinada por el genotipo materno, dado que los factores genéticos
relacionados con el enrollamiento son pasados al embrión por el citoplasma del
ovocito.
Durante el desarrollo, normalmente el primer cuarteto de
micrómeras va a dar origen a la estructura cefálica, del mismo modo el segundo
cuarteto va a contribuir a la formación del estatocisto (órgano del
equilibrio), la concha, el corazón, el velo y la boca; al igual que el segundo
cuarteto, el tercero contribuye a grandes porciones del pie, el velo, el
corazón y además el esófago. Por último la célula 4d (generalmente la de mayor
tamaño), también llamada mesentoblasto está involucrada en la formación del
riñón larval, el corazón, los músculos elevadores y el intestino. La
especificación de estos destinos celulares se da por la ubicación
citoplasmática y por inducción.
Determinación celular:
Los caracoles, al
igual que la mayoría de moluscos, son uno de los mayores ejemplos del
desarrollo en mosaico en las naturales. Durante el proceso de desarrollo, las
blastómeras son especificadas de manera autónoma por localización
citoplasmática, de modo que los determinantes morfo genéticos se encuentran
ubicados en regiones muy específicas del ovocito; es decir, ARN específicos
para algunos factores de transcripción y factores paracrinos se ubican en
células particulares por la asociación con algunos centrosomas. En el caso de
la mayoría de especies de caracoles, éstas moléculas determinantes se
encuentran ubicadas en una región particular del cigoto, la cual va a dar
origen al lóbulo polar.
El lóbulo polar es una estructura que fue descrita por
Wilson y Crampton, los cuales encontraron que durante la segmentación en
espiral de algunos moluscos y anélidos se observaba la extrusión de un bulbo de
citoplasma justo antes de la primera segmentación, este bulbo fue denominado
lóbulo polar. En algunas especies de caracoles, el área de unión entre el
cigoto y el lóbulo se convierte en un tubo estrecho. Durante la primera
segmentación, el cigoto es dividido de forma asimétrica, es por esto que el lóbulo
polar queda únicamente adherido a la blastómera CD. En muchas ocasiones, la
mayor parte del contenido citoplasmático se encuentra almacenado en el lóbulo,
aparentando la presencia de una tercera célula, este estadio es conocido
habitualmente como el estadio de trébol. En el trascurso de la segmentación, el
material del lóbulo polar es absorbido por la blastómera CD y poco tiempo
después, en los momentos previos a la segunda segmentación ocurre una segunda
extrusión. Tras la segunda división, el citoplasma contenido en el lóbulo polar
es absorbido nuevamente, pero esta vez únicamente por la blastómera D.
En 1896,
experimentos realizados por Crampton demostraron que la extirpación del lóbulo
polar durante el estadio de trébol, aunque no afecta la división celular de las
células, ocasiona el desarrollo incompleto de la larva trocófora, esta carece
de estructuras intestinales, corazón y músculos elevadores y ojos entre otros.
Lo anterior demuestra que el citoplasma almacenado en el lóbulo polar contiene
determinantes morfo genéticos involucrados en el desarrollo y diferenciación
del mesodermo y el endodermo. Además explica la contribución de la blastómera D
en la formación del endomesodermo. Con este experimento se concluyó, además,
que los factores de determinación mesodérmica son destinados de manera muy
temprana, poco tiempo después de la fecundación, a tener una localización
específica que determinará el correcto proceso de segmentación de las blastómeras.
Otros estudios sugieren que los determinantes morfo genéticos no se encuentran
ubicados en la parte difusible del citoplasma del lóbulo polar, en cambió
proponen que se localizan en el citoplasma cortical no fluido o en el cito
esqueleto.

Formación del lóbulo porlar durante el desarrollo del caracol
Blastómera
D:
Dado que el material del lóbulo polar es absorbido por la
blastómera D (siendo esta de mayor tamaño), al extirpar esta, o sus blastómeras
derivadas (1D/2D) se va a obtener una larva incompleta similar a la obtenida al
retirar el lóbulo polar. Aunque las blastómera D no contribuyen directamente en
el desarrollo de las estructuras faltantes, parece inducir a otras células a
adquirir los destinos celulares correspondientes. Al retirar la blastómera 3D
inmediatamente tras la formación de 3D y 3d, el fenotipo resultante será
similar a los descritos al eliminar D, 1D o 2D; sin embargo, si la extirpación
se realiza en etapas más tardías la larva resultante es casi completamente
normal. Además se observó que tras la tercera segmentación si se elimina la
blastómera 4D el fenotipo resultante es normal, pero si se retira la célula 4d
la larva carecerá de corazón e intestino. La blastómera 4d es entonces la
responsable en la división subsecuente de dos mesentoblastos (células
precursoras de órganos mesodérmicos (corazón) y endodérmicos (intestino). Por
su parte, la célula 3D induce la activación de la cascada de MAP kinasa en las
micrómeras del ectodermo que se encuentran arriba de ésta y que son precursoras
del ojo y la glándula de la concha.
La polaridad dorso-ventral del embrión, es determinada
también por el material presente en el lóbulo polar. Si se traspasa material
del lóbulo polar a la blastómera AB, además de la blastómera CD, se van a
obtener larvas siamesas unidas en la superficie ventral13
Gastrulación:
La estereoblástula del caracol se caracteriza por ser
relativamente pequeña y tener sus destinos celulares determinados de manera
temprana por la acción de la macrómera D y sus derivadas. La gastrulación
comienza en primer lugar con el proceso de epibolia, de modo que las células
del polo animal se multiplican y cubren las macrómeras del polo vegetal. Por
último, las micrómeras envuelven la totalidad del embrión, dejando una
hendidura de pequeño tamaño en una región del polo vegetal.

Proceso de epibolia durante la gastrulación del caracol
Hibernación:
Hibernación:
Los caracoles hibernan (normalmente desde octubre/abril
hasta finales de abril/octubre). También pueden detener su actividad en verano
en condiciones de sequía, lo que se le conoce como estivación. Para mantenerse
húmedos durante la hibernación, sellan la apertura de su concha con una capa
seca de mucosidad llamada epifragma.
Reproducción:
Los caracoles son hermafroditas, producen tanto
espermatozoides como óvulos. Deben acoplarse porque no pueden auto fecundarse.
Están equipados de un órgano reproductor masculino y del órgano receptivo
correspondiente. Otros, como los caracoles manzana o Ampullariidae, son hembra
o macho.
Los caracoles de jardín, por parejas, se inseminan el uno
al otro, para fertilizar internamente sus óvulos. Generalmente, en la primavera
y el otoño de las zonas templadas, mientras el tiempo permanece caliente y
húmedo. La cópula se hace generalmente de noche y dura de promedio entre 4 y 7
horas. Se lanzan el uno al otro una saeta espiral de carbonato cálcico, que
desaparece en el interior del receptor, donde se disuelve y libera el esperma.
Después hacen un agujero, enterrando sus huevos algunos
centímetros bajo la superficie de la capa fértil. Pasados 15 días, estos huevos
eclosionan y surgen las caracolitas. Cada puesta consiste en hasta 50 huevos.
Son capaces de poner huevos una vez cada mes.

Dos caracoles antes de
la cópula.

Helix
pomatia copulando

Helix
aspersa copulando
Depredadores:
Los caracoles tienen muchos depredadores naturales,
incluyendo otras especies de caracoles, escarabajos, serpientes, sapos,
tortugas, una oruga depredadora y aves como gallináceas, faisánidas, acuáticas,
anátidas, túrdidos, además de un gavilán caracolero especializado en cazar
caracoles de laguna (Ampullariidae).
Se sabe de igual forma que durante su ciclo reproductivo
en la etapa de huevecillos, estos son presa de varios depredadores como el caso
de los ciempiés o escolopendras (quilópodos). Los humanos también aportan
grandes peligros para los caracoles. Además de la amenaza obvia de pisarlos, la
contaminación del agua y la lluvia ácida destruyen sus caparazones y los
envenenan, causando la extinción de muchas de estas especies. Además, los
caracoles son usados en la alimentación humana en todo el mundo.

Conexión del aparato reproductor a la
hora del aparejamiento (Ilustración de Férussac en 1820).
Esperanza de vida:
La vida de los caracoles
varía de una especie a otra. En su hábitat natural, los caracoles Achatinidae
viven alrededor de entre 5 a 7 años y los caracoles del género Helix
aproximadamente de 2 a 3. Los caracoles manzana acuáticos viven sólo 1 año
aproximadamente. La mayor parte de las muertes son debidas a depredadores o
parásitos. En ocasiones, los caracoles han vivido más allá de su vida normal,
hasta los 30 años o más.
Enfermedades:
Los caracoles, tanto
terrestres como acuáticos, son portadores (vectores) de muchos parásitos, tanto
unicelulares como pluricelulares (diversas especies de nematodos) que infectan
a los animales que los ingieren. Se les considera vectores de la gripe.
Hospedadores intermedios de Fasciola gigantica, Fasciola huski, Fasciola
hepatica, trematodos frecuentes en el hígado de los rumiantes. Y en diversas
partes del mundo, como los trópicos, la esquistosomiasis o bilharziosis afecta
a 200 millones de personas.
Frecuentemente utilizando
estrategias inversas al aposematismo los gusanos parásitos delatan al caracol,
para que sea devorado por un predador.
No hay comentarios:
Publicar un comentario